Principle Investigator: Esnart Yohane, Plant Breeder, Department of Agricultural Research Services
Project Objectives: Pigeonpea is an important crop in many parts of Malawi. It is often grown as an intercrop together with maize and can be an important source of protein or additional income for smallholder farmers, can protect the soil from erosion, and can increase biomass production (and improve soil properties) on farm fields. For these reasons, releasing improved varieties (more adapted to Malawi's agroecolgical conditions) can offer smallholder farmers many advantages.
Methods: Ten locally crossed lines of long duration pigeonpea were planted at Chitedze Agricultural Research Station. Crops were planted at the beginning of the rainy season in January 2015. On May 28 and June 12 (after flowering but before pod-fill), the MultispeQ was used to measure relative chlorophyll content (SPAD) and the quantum yields of photosystem 2 (Phi 2), non-photochemical quenching (PhiNPQ) and other energy losses (PhiNO). The PhotosynQ protocols used to measure these parameters were ‘Chlorophyll content (SPAD) I’ and ‘The One Protocol (Phi2, PSI, NPQ) II.’ In each plot, leaves from three different canopy positions--upper, middle, and lower--were measured from 6 plants. At physiological maturity, pigeonpea was harvested and seed weight (per 100 seeds) and pigeonpea yield were calculated for each plot.
The study was a complete randomized block design with 3 replicates. Initial analysis was completed using a one-way ANOVA. Multiple linear regression analysis was then used to determine the effects of light intensity, time of day when the leaf was measured, leaf position in the canopy, and pigeonpea variety on SPAD, ΦII, ΦNPQ, and ΦNO and effects were considered significant at p ≤ 0.05. Mean separation of varieties was based on Tukey’s honest significant difference (HSD) test and differences were considered significant at p ≤ 0.05. A mixed effects model, with light intensity, time of day, and leaf position as fixed effects and block has a random effect was used to generate unique coefficients for each plot. Simple regression was used to test for correlations between these unique coefficients and crop yield and seed size. This model was run to include all leaves from all three leaf positions and for each leaf position independently. Effects were considered significant at p ≤ 0.05.

Results: First, we analyzed each variety using gross production parameters (i.e. crop yield and seed size) and simple aggregate PhotosynQ parameters (Table 1). Using this method their were no significant varietal differences in either gross production parameters or PhotosynQ parameters. The lack of significant differences between different varieties can most likely be attributed to: 1) large standard deviations in gross production parameters and 2) the effect of potentially influencing factors such as light intensity and time of measurement on photosynthesis parameters.
Next, we accounted for factors affecting photosynthesis (i.e. time of day when the measurement was taken, light intensity, block, and leaf position on the plant) with multiple linear regression. PhotosynQ data provided useful insights into different varieties ability to capture and use light energy (Figure 1). The variety ICEAP 01535 had significantly higher relative chlorophyll content than ICEAP 01494, ICEAP 01499, and ICEAP 01198 and significantly higher ΦII than ICEAP 01198. Also, the variety ICEAP 01527 had significantly higher ΦII than ICEAP 01499 and ICEAP 01198. These results suggest that these two varieties were efficient at capturing and using light energy, while ICEAP 01494, ICEAP 01499, and ICEAP 01198 were the worst varieties at capturing and using light energy. However, these results do not explain how that energy was used (i.e. produce grain, vegetative growth, etc).

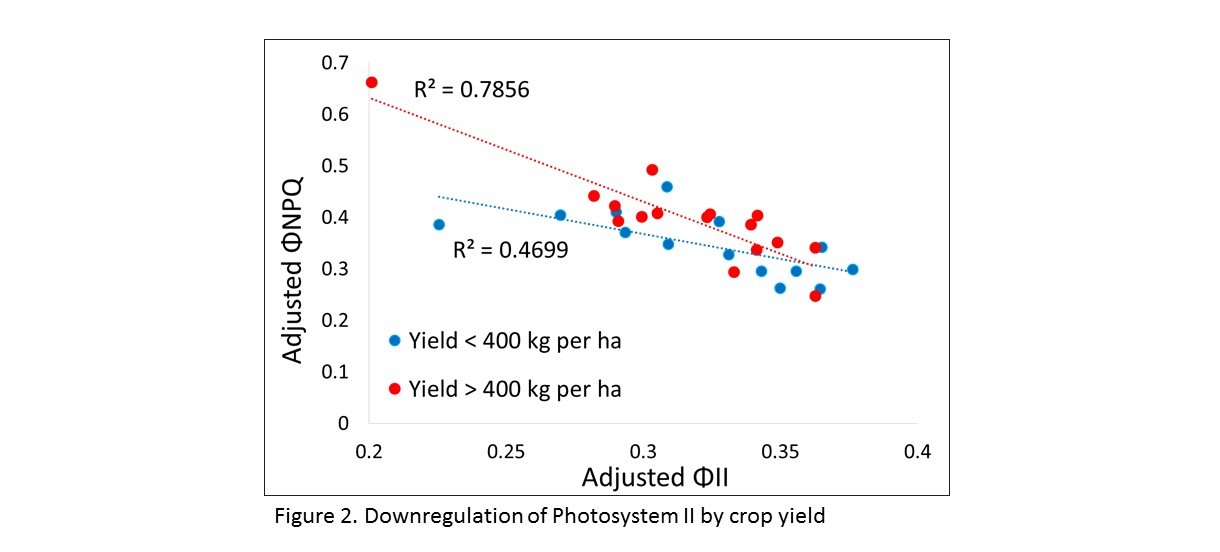
When combining all leaves into the mixed effects and simple regression models, there was a significantly positive correlation between ΦNPQ and yield and negative correlation between ΦNO and yield (Table 2). Analyzing only the top leaves resulted in a similar relationship between PhotosynQ data and crop yield. The interactions between ΦNPQ and ΦNO represent the ability of plants to either regulate away excess light energy or endure photo-damage. For this reason, the directions of the relationships between ΦNPQ and ΦNO and yield are inverse. To better understand the effect of downregulation of Photosystem II on yield, we graphed the relationship between ΦII and ΦNPQ for high and low yielding pigeonpea (Figure 2.). The higher yielding pigeonpea (regardless of variety) had a greater regulatory response to reductions in ΦII than low yielding pigeonpea. When analyzing only the middle or bottom leaves, there was no relationship between yield and PhotosynQ data. Instead, SPAD was negatively correlated with seed size for both the middle and bottom leaves.

It appears the plants that were better able to regulate photosynthesis (ΦNPQ) had higher crop yields. Based on both Photosynthetic measurements and plant growth outputs, the variety ICEAP 1527 appears to be the best performing variety for the agroecological conditions at Chitedze Research Station. High variability and a limited number of PhotosynQ measurements (data was only collected late in the season) limited the ability to conduct a robust analysis of the data. However, the data collected still provide valuable information to plant breeders. In the future, it would be better to collect data more often (and at earlier growth stages) and to collect data from more than one site.